1) Neutron moisture meter

中性子水分儀
2) Neutron moisture meter
中性子水分仪
3) Neutral molecule
中性分子
1.
Microwave assisted synthesis of chiral molecular tweezers based on deoxycholic acid and its recognition properties for neutral molecules;
脱氧胆酸手性分子钳的微波合成及其对中性分子的识别性能研究
4) neutral molecules
中性分子
1.
The basic concepts and principles, experimental schemes and results as well as recent progress of manipulation and control of cold neutral molecules and their geometric molecule-optical devices in the world are introduced.
主要介绍了国际上有关中性分子操纵与控制及其几何分子光学器件的基本概念、基本原理、研究方案与结果及其最新进展。
5) Liquid limit apparatus
液性限度儀
补充资料:分子进化的中性学说
| 分子进化的中性学说 molecular evolution,neutral theory of 认为分子水平上的大多数突变是中性或近中性的学说。自然选择对它们不起作用,这些突变全靠一代又一代的随机漂变而被保存或趋于消失,从而形成分子水平上的进化性变化或种内变异。简称中性学说。这一学说由日本遗传学家木村资生于1968年首先提出。 提出中性学说的背景 20世纪50年代以后,先后搞清楚了许多生物大分子的一级结构。通过比较不同生物的某些功能相同的蛋白质或核酸的氨基酸或核苷酸序列的差异,人们发现,亲缘关系近的差异较小,亲缘关系远的差异较大,与物种的表型进化情况基本一致(见分子进化)。分子进化有3个特点。一是多样性程度高,与表型多态(即在一相互交配的群体中存在着两种或多种基因型的现象)相比,分子多态更为丰富(例如细胞色素C这种蛋白质分子在行有氧呼吸的不同物种中就有种种不同的分子结构)。二是各种同源分子能很好地完成各自的功能(如脊椎动物的血红蛋白分子都能运氧、各种生物的细胞色素C都能在氧化磷酸化中完成电子的传递等)。三是随着生物从低级向高级演化,同源分子中逐年发生氨基酸或核苷酸的替换,且大致按每年每位置替换数恒定速率进化。关于大分子的进化性变化,早在1965年E. 楚克坎德尔和L.C.波林等就已有过全面的论述。木村的功绩是在理论上更迈进一步,把中性突变-遗传随机漂变放到决定性的位置上,提出分子进化的中性学说,较合理地解释了分子进化的各种现象。 中性学说的依据 中性学说的出发点是中性突变。DNA产生突变的方式有4种,即核苷酸的替换、缺失、附加和倒位,而最常发生的是核苷酸的替换。对一个密码子来说,由于核苷酸替换可发生在三联体的任一位置上,而每一位置都可能有3种替换方式,故一个核苷酸替换可有9种方式导致密码子变换,如AAA→AA 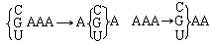 因此,61种氨基酸密码就可因一个核苷酸替换而产生(61×9=)549 种密码子变换。AAA与AAG都是Lys的密码子,这种密码子变换并不引起氨基酸替换,称同义突变或沉默突变。AGA是Arg的密码子,UAA是终止密码子,从AAA到AGA引起氨基酸替换,称为变义突变;从AAA到UAA则称无义突变,因为UAA代表终止信号 。如果将549种变换对照遗传密码表逐一统计,可以发现由于遗传密码的简并有134/549=24.4%即约1/4的替换为同义突变。 因此,61种氨基酸密码就可因一个核苷酸替换而产生(61×9=)549 种密码子变换。AAA与AAG都是Lys的密码子,这种密码子变换并不引起氨基酸替换,称同义突变或沉默突变。AGA是Arg的密码子,UAA是终止密码子,从AAA到AGA引起氨基酸替换,称为变义突变;从AAA到UAA则称无义突变,因为UAA代表终止信号 。如果将549种变换对照遗传密码表逐一统计,可以发现由于遗传密码的简并有134/549=24.4%即约1/4的替换为同义突变。氨基酸替换又分为不影响蛋白质功能的、性质相似的氨基酸之间的替换即保守替换(如Arg和Lys间,Phe和Tyr间,Glu和Asp间等)和影响蛋白质功能的、性质明显不同的氨基酸之间的替换即非保守替换(如Asp和Val间,Phe和Ser间,Arg和Ile间等)。若每种方式以同样概率发生,则由一个核苷酸替换所引起的全部氨基酸替换中有53%是保守替换,而从分子进化研究中观察到的氨基酸替换来分析,保守替换可高达68%。同义突变和保守替换不改变或很少改变分子的结构与功能,所以在选择上是中性的或近中性的。此外,DNA的非编码区以及基因的内含子区所发生的核苷酸替换,以及不转译成蛋白质的假基因中所发生的变化等,也是中性或近中性的。自然选择对中性或近中性的突变不起作用,在众多的中性突变中,哪一种变异能够流传,哪一种变异趋于消失,全靠机遇,这种基因频率的随机变动就是遗传漂变。遗传漂变最初由S.赖特于20世纪30年代提出。他认为,当一个大群体划分为若干个小群体繁殖时,群体的基因组成实际上等于进行了一次随机取样。 赖特认为遗传漂变仅发生在小群体中。木村则证明,遗传漂变并不限于小群体,对任何一个大小一定的群体,都能通过遗传漂变引起基因的固定,从而导致发生进化性变化 。 分子进化速率与中性学说 分子进化速率是以每年每位置氨基酸或核苷酸替换数来表示的。以蛋白质分子为例,其进化速率为: 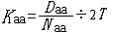 (1)其中Daa是两种不同生物同源蛋白质的氨基酸差异数,Naa是构成同源蛋白质的氨基酸个数,T是两种生物的分歧进化时间。(1)式中之所以要除以2T,是因为分歧进化是向两条路线进行的。可以这样理解:从一种生物回溯到分歧点再到第二种生物,其演化时间正好是2T。分子进化的特点之一是每一种生物大分子不论在何种生物都有一个大致恒定的进化速率。以血红蛋白α-链为例,鲤与马有66个氨基酸差异,地质资料表明鱼类约起源于4亿多年前的志留纪,若以4亿年作为鱼与马的分歧进化时间,则从鱼到马的进化速率为: 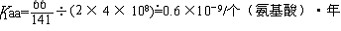 马与人有18个氨基酸差异,高等哺乳动物的辐射进化约出现于8000万年前,如以此作为马与人的分歧进化时间,则从马到人的进化速率为: 马与人有18个氨基酸差异,高等哺乳动物的辐射进化约出现于8000万年前,如以此作为马与人的分歧进化时间,则从马到人的进化速率为: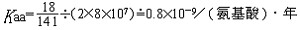 即血红蛋白的α链分子,无论是在从鱼到马还是在从马到人的进化过程中,其进化速率基本上都是相同的。分子进化速率与种群大小、世代寿命和物种的生殖力均无关,也不受环境因素所影响。这是自然选择学说所不能解释的,而中性学说对此却能作出既简明又合理的解释。 即血红蛋白的α链分子,无论是在从鱼到马还是在从马到人的进化过程中,其进化速率基本上都是相同的。分子进化速率与种群大小、世代寿命和物种的生殖力均无关,也不受环境因素所影响。这是自然选择学说所不能解释的,而中性学说对此却能作出既简明又合理的解释。设大小为N的一个二倍体群体中,共有2N个基因位点 ,若每配子每代的突变率为v,则每代将产生2Nv个新突变。设一个突变最终被固定的概率为μ,则每代每位点的突变替换数(进化速率)为: K=2Nvμ (2)由于这个群体中有 2N个基因,当突变呈中性时,每个基因被固定的机会都相同 ,即 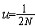 。于是2Nv个突变中仅有1/2N个被固定,即: 。于是2Nv个突变中仅有1/2N个被固定,即: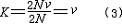 可见,在中性突变情况下,进化速率是一个恒定值且等于其突变率。 可见,在中性突变情况下,进化速率是一个恒定值且等于其突变率。中性学说只是强调分子水平上多数突变是中性的,并没有说全部突变都是中性的。对于蛋白质来说,只有不改变分子的三级结构和功能的那些氨基酸替换,才大致保持每年每位置上的恒定速度,否则就要受到自然选择的作用。如血红蛋白-β链上的大多数氨基酸替换都是中性替换,这些替换的固定都是通过遗传漂变实现的;但如果其第 6位的谷氨酸被缬氨酸所替换,则在人血中就会出现不能运氧的镰形红细胞。如果所有红细胞都是镰形细胞(纯合体),个体未到成年就会死去,这样的突变当然要受到自然选择的作用而最终被淘汰;而当基因处于杂合状态时,由于杂合体(只有部分红细胞为镰形细胞)提高了抗疟能力,所以在某些疟疾流行的地区这种杂合体就有明显的优势,以致此突变基因在该地区有非常高的频率且能保持稳定。此种情况完全可用综合进化论的超显性说加以说明,而不能用遗传漂变来解释。生物大分子虽然有丰富的多样性,但要维持其正常的结构与功能,其替换就不可能完全随机;不同分子受到的限制不同,所受到的选择清除程度也不一样。由此可见,中性学说和达尔文的自然选择学说不是对立的。不少学者认为,在理解生物进化上可把中性学说看作是附加在自然选择学说中的一个原理,是在分子水平上对达尔文主义的补充和发展。 |
说明:补充资料仅用于学习参考,请勿用于其它任何用途。
参考词条